
国土数値情報の洪水浸水想定区域データをQGISで表現するではバッチプロセスで属性テーブルの属性をファイル名に取り出す部分でつまづいていた.今回QGISのコミュニティで質問した結果回答が得られたので結果を公開する.
“国土数値情報の想定最大規模の洪水浸水想定区域を浸水深ランクコードごとにQGISのバッチプロセスで融合する” の続きを読む
国土数値情報の洪水浸水想定区域データをQGISで表現するではバッチプロセスで属性テーブルの属性をファイル名に取り出す部分でつまづいていた.今回QGISのコミュニティで質問した結果回答が得られたので結果を公開する.
“国土数値情報の想定最大規模の洪水浸水想定区域を浸水深ランクコードごとにQGISのバッチプロセスで融合する” の続きを読む
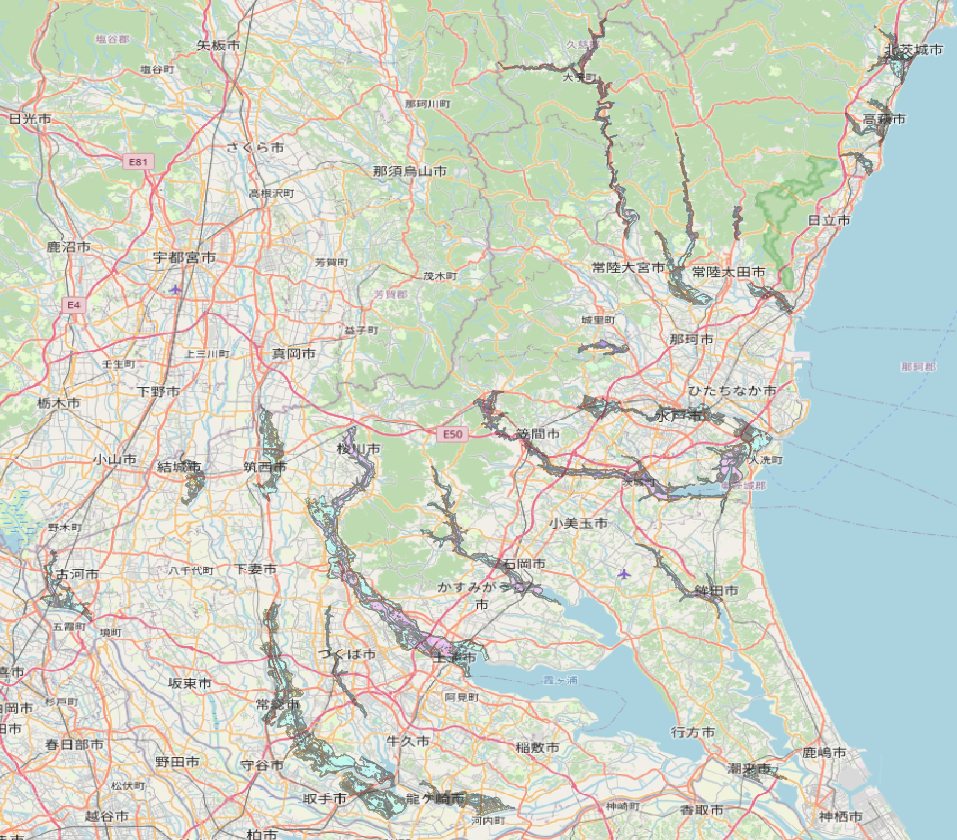
この記事が公開される頃には梅雨真っ盛りであり,各地で洪水の被害が報道されていることと思う.いささか遅きに失した感もあるが,令和5年度版の洪水浸水想定区域が5月31日に発表されたため,記事にした.とりあえず茨城県版のデータをQGISに取り込み,浸水深ランクコードごとに塗り分けた.各自治台の担当に置かれては洪水ハザードマップの更新に忙しい時期と思われ,参考にされたい.
温暖化に伴い洪水被害が増加しつつある.各自治体のハザードマップの重要性が増しているところであるが,今回は国土数値情報ダウンロードサイトから洪水浸水想定区域のデータをダウンロードし,QGISで表現してみた.
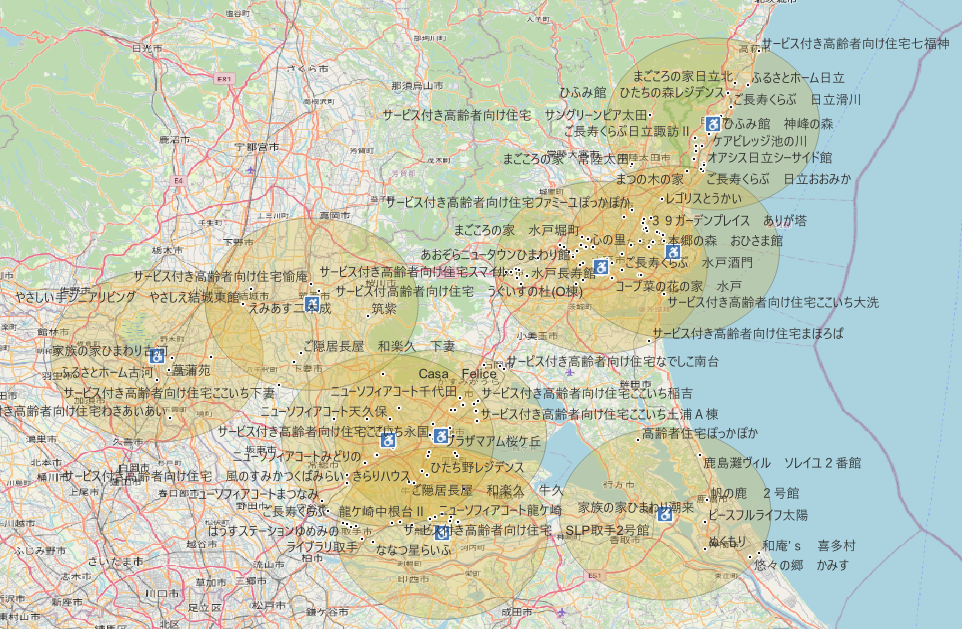
今回は保健所から16km圏内にあるサービス付き高齢者住宅を抽出し,QGISで表現したい.基準となる地点は何でも良い.国土数値情報ダウンロードサイトにあるデータをダウンロードし,QGISで読み込む形である.
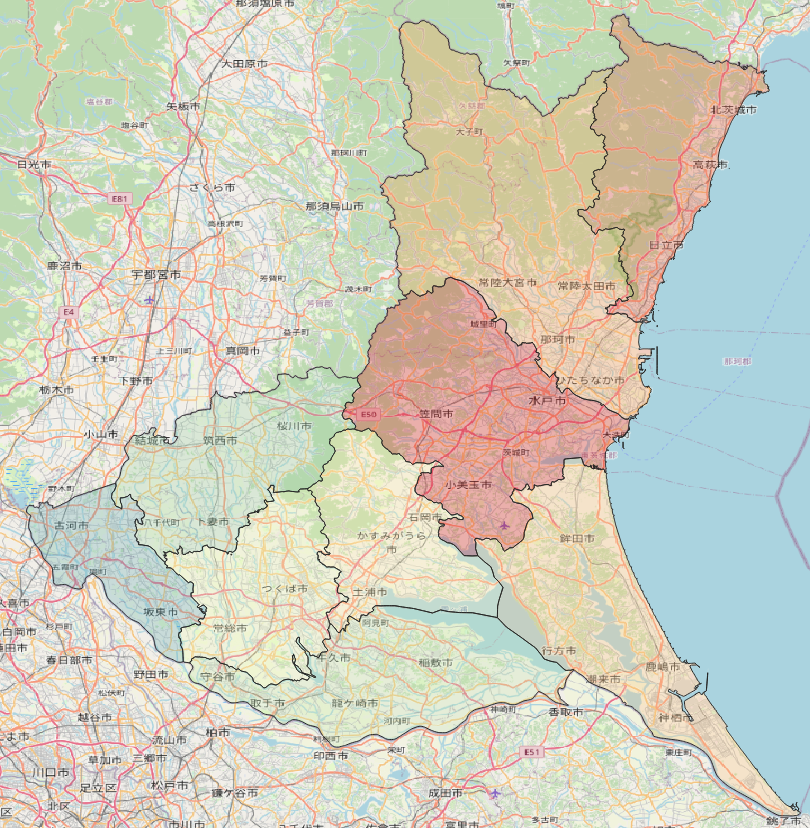
QGISシリーズである.今回は茨城県の二次医療圏をカラーサンプルを使って塗り分ける.
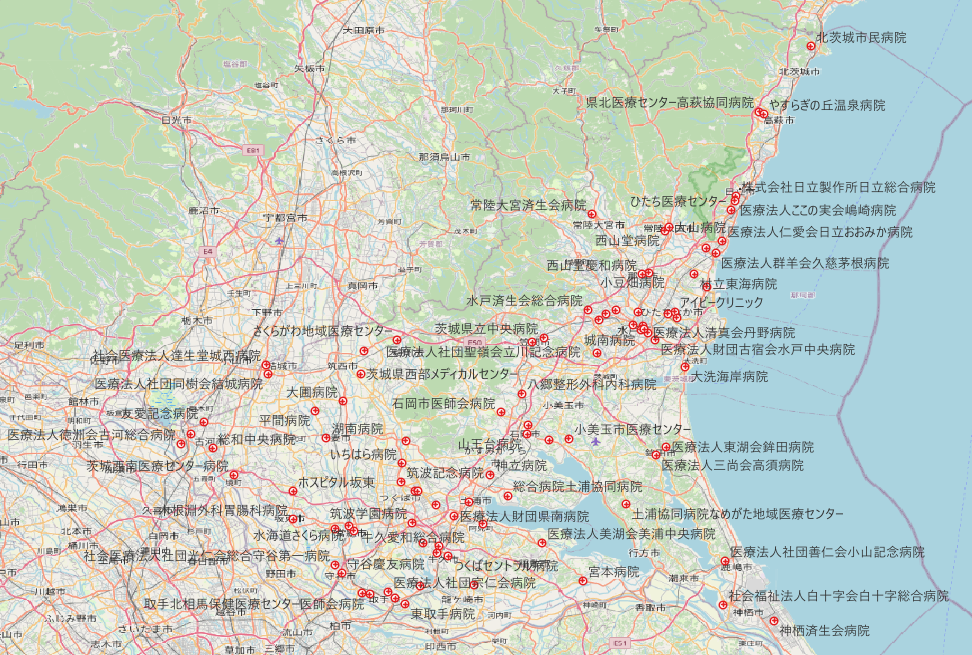
国土数値情報ダウンロードサイトには様々なデータが蓄積されている.医療機関の位置データもその一つである.今回は茨城県の救急告示病院をQGISで表現することにした.
前回の投稿(心血管系または呼吸器系の入院歴のある高齢コホートメンバーの死亡リスクに及ぼす断熱改修および暖房の影響)では循環器系疾患および呼吸器系疾患による死亡率が断熱介入により減少することをみた.今回は断熱改修の費用対効果の経済的側面に焦点を当てる.
原著はここから読める.健康上の利益とエネルギー上の利益は,コストを2倍近く上回る,というのが本著の趣旨である.日本においてもこのようなエビデンスに基づく政策が実行されてほしい.
先日筆者は新型コロナウイルスに感染した.その際,自宅での感染隔離対策が読者にとって何か示唆する所があればと思い記事にすることにした.
この記事がすべての住宅に当てはまるわけではない.住宅の構造に帰するところが大きい点もある.家族構成によっては当てはまらない点もある.それを踏まえて読んでほしい.
二乗平均平方根誤差とは英語では Root Mean Squared Error (RMSE) と書く.真値と予測値との乖離(誤差)を二乗し,その平均値をとり,その平方根を求めた値のことである.非負の値を取り,0に近いほど優れたモデルであることを示唆する.
今回使用するのはe-Statからの社会疫学的指標を加えて熱中症搬送人員数を分析するで使用したデータベースである.先の記事では回帰モデルを評価する指標が必要との認識であった.